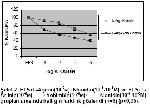
Çalışmamızda izole sıçan ileum preparatlarında a2-
agonisti klonidinin efektif dozunu belirlemek amacıyla
ortama kümülatif konsantrasyonlarda ilave edilmiştir.
Ortama klonidin ilave edildiği zaman kontrol yanıtları
olarak kabul ettiğimiz EFS yanıtlarına göre kasılma
yanıtlarında doza bağımlı bir azalma meydana geldi.
Klonidininin 10-8 derişiminde elde edilen kasılma
yanıtları % 87±2,7, 10-7 derişimde % 75,8±4,2, 10-6
derişimde % 67,3±3,7, 10-5 derişimde ise % 55,3±1,8
olarak değişmiştir (n=6) (p<0,05) (Şekil 1). Ortama a2- adrenoseptör antagonisti yohimbinin ilave edilmesi ile
klonidininin 10-8 derişiminde elde edilen kasılma yanıtları
% 103,6±4,9, 10-7 derişimde % 96,4±5,3, 10-6 derişimde
% 87,4±4,1, 10-5 derişimde ise % 81,7±5,2 olarak
gözlenmiştir. Klonidin tarafından oluşturulan bu
inhibisyon yohimbinin tarafından baskılanmıştır (n=6)
(p<0,05) (Şekil 1).

Büyütmek İçin Tıklayın |
Şekil 1: Klonidin (10-8-10-5M) ve Yohimbin (10-6м)+
Klonidin(10-8-10-5M) grupları arasında belirgin farklılık
gözlenmiştir (n=6) (p>0,05). |
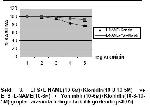
Nitrik oksit aracılı gevşeme yanıtları üzerine olası
katkısının incelenmesi amacıyla ortama ilave edilen NO
prekürsörü L-Arg, klonidinin EFS ile indükte kasılma
yanıtları üzerine olan inhibitör etkinliğini artırdığı
gözlenmiştir. Klonidininin 10-8 derişiminde elde edilen
kasılma yanıtları % 68,6±1,3, 10-7 derişimde %
57,3±3,1, 10-6 derişimde % 48±0,7, 10-5 derişimde ise
% 40,7±2,6 olarak değişmiştir (n=6) (p<0,05) (Şekil 2) ve
benzer olarak kurgulanmış farklı seride ortama yohimbin
ilave edilmesi durumunda kasılma yanıtlarının anlamlı
olarak azaldığı gözlenmiştir. Bu şartlarda klonidinin 10-8
derişiminde elde edilen kasılma yanıtları % 92,5±3,7, 10-
7 derişimde % 86,9±4,2, 10-6 derişimde % 70,7±3,5, 10-
5 derişimde ise % 56,8±2,3 olarak değişmiştir (n=6)
(p<0,05)(Şekil 2). Kasılma yanıtları üzerine inhibitör
etkinliğinin sadece L-Arg bulundurulan gruba göre
anlamlı şekilde azaldığı görüldü (p<0,05)(Şekil 2).
Büyütmek İçin Tıklayın |
Şekil 2: EFS+L-Arjinin(10-6м)+Klonidin(10-8-10-5M) ve EFS+LArjinin(
10-6м) +Yohimbin(10-6м)+ Klonidin(10-8-10-5M)
grupları arasında belirgin farklılık gözlendi (n=6) (p<0,05). |
Nitrik oksit sentaz inhibitörü L-NAME bulundurulduğu
ortama ilave edilen klonidin kümülatif dozlarında EFS'a
verilen kasılma yanıtları klonidinin 10-8 derişiminde % 100,7±2,4, 10-7 derişimde % 96,2±2,1, 10-6 derişimde %
83,4±3,9, 10-5 derişimde ise % 77,5±1,2 olarak
değişmiştir ancak bu yanıtlar arasında anlamlı bir farklılık
gözlenmemiştir (n=6)(p>0,05)(Şekil 3). Ortamda
yohimbin ve L-NAME'in birlikte bulundurulduğu halde
kasılma yanıtları klonidinin 10-8 derişiminde % 100±4,7,
10-7 derişimde % 99,4±3,2, 10-6 derişimde % 96,1±2,7,
10-5 derişimde ise % 93,1±4,1 olarak değişmiş ve
yanıtlar arasında anlamlı farklılık gözlenmiştir (n=6)
(p<0,05)(Şekil 3). Ortamda yohimbin bulundurulduğu
halde EFS kasılma yanıtları üzerine inhibitör etkinliğinin
sadece L-NAME bulundurulan gruba göre anlamlı şekilde
azaldığı görülmüştür (p<0,05)(Şekil 3).
Büyütmek İçin Tıklayın |
Şekil 3: EFS+L-NAME(10-6м)+Klonidin(10-8-10-5M) ve
EFS+L-NAME(10-6м) + Yohimbin(10-6м)+Klonidin(10-8-10-5M) grupları arasında belirgin farklılık gözlendi (p<0,05). |
Klonidinin EFS'a verdiği kasılma yanıtları ortama LArg
ilave edildiği durumda belirgin olarak azalırken
(p<0,05)(Şekil 4), L-NAME bulundurulan seride anlamlı
bir değişiklik gözlenmemiştir (p<0,05)(Şekil 4).

Büyütmek İçin Tıklayın |
Şekil 4: EFS+Klonidin (10-8-10-5M), EFS+ L-Arjinin (10-6 м)+
Klonidin (10-8-10-5M), EFS+L-NAME(10-6 м)+Klonidin (10-8-10-5M) grupları arasında belirgin farklılık gözlenmiştir (n=6)
(p<0,05). |
Klonidinin ortamda yohimbin bulunduğu halde EFS'a
verdiği kasılma yanıtları farklı serilerde ortamda L-Arg
bulundurulduğunda belirgin bir inhibisyona neden olurken
ortamda L-NAME bulunduğu halde anlamlı bir farklılık ile
karşılaşılmamıştır (p>0,05)(Şekil 5). Ancak ortamda LArg
ve yohimbin bulundurulduğu durumda klonidinin
kümülatif derişimlerinde kasılma yanıtlarında inhibisyon
görülürken, L-NAME+ yohimbin bulundurulduğu
durumlarda kasılma yanıtları üzerine olan inhibisyonda düşüş kaydedilmiştir (p<0,05)(Şekil 5). Bu üç grup
karşılaştırıldığı zaman aralarında anlamlı farklılık ile
karşılaşılmıştır(p<0,05)(Şekil 5).

Büyütmek İçin Tıklayın |
Şekil 5: Yohimbin(10-6м)+Klonidin(10-8-10-5M), L-Arg(10-6м)+Yohimbin(10-6 м)+Klonidin (10-8-10-5M) ve L-NAME(10-6
м)+Yohimbin(10-6 м)+Klonidin(10-8-10-5M) grupları kasılma
yanıtları arasında anlamlı farklılıklar gözlendi (n=6) (p<0,05). |
Elektriksel alan stimülasyonu ile elde edilen kasılma
yanıtları üzerine klonidinin inhibitör etki gücünü
göstermek amacıyla saptanan IC50 değerleri aşağıda
gösterilmiştir (Tablo 2).

Büyütmek İçin Tıklayın |
Tablo 2: Gruplar arasında IC50 değerleri açısından farklılık gözlendi. Gruplar ve ortalama -log IC50 değerleri (n=6) (p<0,05). |


)
)
)
)
)
)
)
